There are many physical factors that effect fish bioenergetics, such as local hydrologic characteristics in a stream or river. However, not all of the physical factors involved are especially obvious. We will look at three examples here.
Water velocity plays an extremly important role in fish bioenergetics. It effects components that are detrimental to fish as well as components that are beneficial. The downstream force of water is something that the fish must expend energy against unless it wants to be washed away. As pointed out on the previous page, a fish needs to exert a force equal and opposite to the downstream force of water in order to maintain a stationary position. In order to move upstream or make lateral moves, it needs to exert a force greater than the downstream force of water. While water velocity demands energy from the fish, it also effects how much energy is available for the fish to gain. As shown in the graph below, for a given amount of drifting invertebrate energy in a stream, there is an increase in the amount availlable to the fish with an increase in water velocity. Therefore, water velocity will variably effect a fish, depending on the amount of energy available.
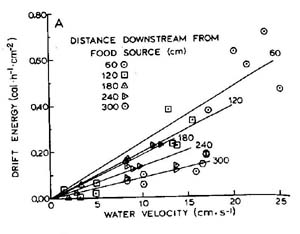
Figure from Fausch (1984)
Because water velocity is such an important component of fish bioenergetics, stream characteristics that effect water velocity can strongly influence where a fish hangs out. For example, structural components such as large woody debris that cause changes in local hydrologic characteristics are highly beneficial to fish. An extremely profitable position for a fish occurs where the fish can spend most of its time in slower moving water that is immediately adjacent to faster water. Staying in slower water reduces the energetic cost to the fish while the faster water next to it provides a more benficial (higher) rate of energy in the form of drifting invertebrates. The fish can hold its primary position in the slow water and move into the faster water to retrieve prey for brief periods. As shown in the experimental results below, most fish will choose the highly beneficial positions where there is physical structure. A total of 113 juvenile coho salmon selected a structured habitat compared to many fewer that were found in unstructured habitats of both slow and fast water.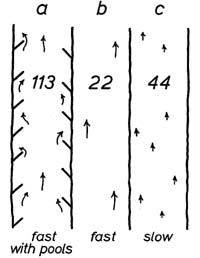
Figure from Mundie (1969)
There are also some important forces operating on a microscale. When a fish is feeding on drifting macroinvertebrates or zooplankton, the amount of prey that it can take into its mouth at one time is a function of the volume that is inhlaled through the mouth and gills by expanding and then contracting these parts. Small, mobile invertebrates on the periphery of this volume are able to escape sometimes when their swimming strength is greater than the inhaling force. The speed with which these invertebrates must move in order to evade going into the fish’s mouth is the escape velocity. The retention of prey that does enter the mouth is largely a function of gill-raker spacing, with organisms that are smaller than these spacings often passing through the gills unscathed. However, a healthy fish will also have a signifiacnt layer of mucous on the gills so that some of the smaller prey items can be retained. Many small organisms can be caught in the mucous when the strength of the adherence is greater than the force of the water pushing through the gills, much like static friction will keep things stationary. The net result of these various micro-components will determine how much energy is acquired by the fish. (figure, Wankowski 1979).
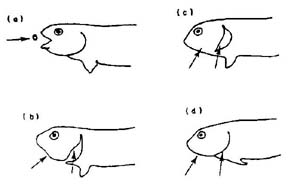
Figure from Wankowski (1979)